








YK11 SARM POWDER – 1000MG / 1 GRAM


$70.99
YK-11 SARM is sold for laboratory research use only. Terms of sale apply. Not for human consumption, nor medical, veterinary, or household uses. Please familiarize yourself with our Terms & Conditions prior to ordering.


*Includes:
- One (~10mg – 15mg) Red Micro Scoop
Also Available In:
![]() Liquid Option >>
Liquid Option >>
![]() Gel Option >>
Gel Option >>
- Description
- Additional information
Description
YK11 SARM Powder
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
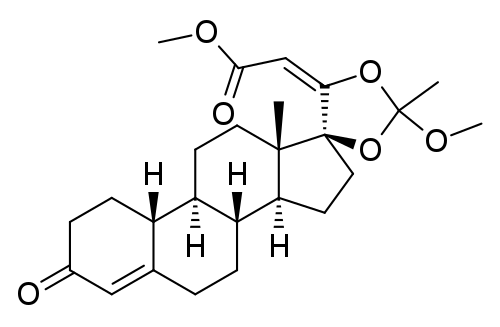
| CAS Number | 1370003-76-1 |
| Other Names | YK11, YK 11, Z9748J6B0R, Z9748J6B0R, UNII-Z9748J6B0R, EX-A727, DTXSID301107018, VEC00376 |
| IUPAC Name | methyl (2E)-2-[(8R,9S,10R,13S,14S,17S)-2′-methoxy-2′,13-dimethyl-3-oxospiro[1,2,6,7,8,9,10,11,12,14,15,16-dodecahydrocyclopenta[a]phenanthrene-17,5′-1,3-dioxolane]-4′-ylidene]acetate |
| Molecular Formula | C₂₅H₃₄O₆ |
| Molecular Weight | 430.5 |
| Purity | ≥99% Pure (LC-MS) |
| Liquid Availability |
|
| Powder Availability | |
| Gel Availability | |
| Storage | Store in cool dry environment, away from direct sunlight. |
| Terms | All products are for laboratory developmental research USE ONLY. Products are not for human consumption. |
*Includes:
- One (~10mg – 15mg) Red Micro Scoop

What is YK11?
(17α,20E)-17,20-[(1-methoxyethylidene)bis(oxy)]-3-oxo-19-norpregna-4,20-diene-21-carboxylic acid methyl ester, is a non-steroidal selective androgen receptor modulator (SARM), more commonly referred to as YK11. YK11 has shown potential in effectively increasing bone density and muscle mass while reducing fat mass. Studies have suggested that YK11 is able to improve osteoblast activity and bone density through activation of the protein kinase B.That being said, current research indicates that muscle growth caused by YK11 is due to increased levels of follistatin. Increased levels of follistatin lead to the inhibition of myostatin, a protein responsible for the regulation of muscle growth. With myostatin blocked as a result of YK11 treatment, there is no inhibition of potential muscle growth. The SARM is capable of eliciting responses in the anabolic pathways that are similar to those associated with traditional anabolic agents, testosterone and DHT.
Main Research Findings
1) The SARM, YK11 has shown the potential to successfully act as a partial agonist to the androgen receptor.
2) Proliferation and differentiation of osteoblasts in MC3T3-E1 mouse cells is enhanced through supplementation of YK11.
Selected Data
1) The research team of Kanno et. Al conducted an experiment in order to thoroughly examine the agonistic properties of YK11. The study began by preparing a 100 mL two-necked round-bottomed flask by adding a magnetic stirring bar, 6.5 mg, 0.025 mmol of (CH3CN2)PdCl2, 108.1 mg, 1.0 mmol of p-benzoquinone, and 7 mL of MeO to the flask. It was then fitted with a rubber septum and three-way stopcock connected to a balloon filled with carbon monoxide (CO). A 170.3 mg/ 0.5 mmol solution of substrate 1 in 35 mL of MeOH was added dropwisely to the stirred mixture. The mixture was stirred for 70 hours, followed by additional dilution with 70 mL of CH2Cl2, washing with 50 mL of 5% aq NaOH, drying over MgSO4, and concentration under reduced pressure. The product was purified through the use of column chromatography and the resulting fraction was eluted with hexane/ethyl acetate (15/1) solution. The purification process yielded YK11 (93 mg, 43%) as a mixture of inseparable diastereomers [1].
Following the purification process , various tests were performed in order to assess how YK11 acts as an agonist to androgen receptors (AR). Cell culturing of the human embryonic kidney cell in, HEK293, and the human breast cancer cell line, MDA-MB 453 expressing wild-type AR, took place first. Culturing of the cells took place in Dulbecco’s modified Eagle’s medium containing 10% fetal bovine serum (FBS) and penicillin-streptomycin in a humidified climate containing 5% CO2. The HEK293 cells were transfected with pcDNA5/TO-AR. The 100 mcg/mL of hygromycin medium was replaced the next day and all sub-cultured colonies were collected by the researchers after 2 weeks of treatment. The androgen receptors that expressed HEK293 cells were then labeled as the 293AR cell line [1].
The researchers conducted a luciferase reporter analysis after cell culturing. Cells were seeded in 48-well plates and transfected with various expression plasmids. The ARE-luciferase reporter plasmid and a Renilla pGL4.74 [hRluc/TK] were used as the internal standards of the study as it was determined through the reverse-transfection method using a transfection reagent. The cells were incubated overnight in phenol DMEM containing 5% charcoal-stripped FBS, followed by 24 hours of AR ligand treatment. Luciferase activity was able to be measured through the use of Dual-Luciferase Reporter Assay System and a Turner Designs Luminometer.
The next step of the study was to complete mammalian two-hybrid assays. Similar to the luciferase assay, the cells were seeded in 48 well plates and transfected. However, these cells were transfected with the expression plasmids encoding the GAL4 and VP16 fusion proteins, the pG5-Luc reporter plasmid, and pGL4.74. Transfection occurred through the reverse transfection method that utilizes the GeneJuice transfection reagent. The cells were then incubated overnight in phenol red-free DMEM containing %5 csFBS, followed by AR ligand treatment for 24 hours [1].
The GeneJuice transfection agent was also used in combination with 500 ng of Yellow fluorescent protein (YFP)-tagged AR in the transfection process of HEK293 cells. This process was performed on 4-well chambered coverglasses, followed by incubation overnight and treatment with either YK11 or DHT over a 6 hour time period. Localization of YFP-tagged proteins was observed with a Zeiss LSM 510 confocal laser scanning microscope. The researchers then completed a real time reverse transcription (RT)-PCR in order to detect the genetic makeup of the cell culture samples. The cells were seeded in 6-well plates in a medium of phenol red-free DMEM containing 5% csFBS. The plates were left overnight and treated with either DHT or YK11 over a 24 hour treatment period [1].
Following treatment with YK11 or DHT, TaKaRa RNAiso Reagent was used to isolate total RNA, which was then digested with DNase1. Synthesization of cDNA took place with the Rever Tra Ace qPCR RT Kit, while the RT-PCR was performed with the SYBR Green Real-time PCR Master Mix-plus, in addition to PCR primers composed of varying oligonucleotide sequences. The final test conducted by the research team was a Western blot analysis. Sodium dodecyl sulfate (SDS) was used to extract the total protein. Total protein was further supplemented with a potent protease inhibitor cocktail, while the protein concentration was then measured using a 2-D Quant kit. The protein samples were separated through the use of SDS-polyacrylamide gel electrophoresis (SDS-PAGE).
Western blot analysis was performed on the cell cultures using rabbit anti-AR antibody as the primary antibody, and horseradish peroxidase-conjugated anti-rabbit immunoglobulin as the secondary antibody. Using the Immobilon Western Detection Reagent, researchers were able to effectively measure the protein bands present [1].
2) The research team of Yatsu et. Al examined the potential of YK11 to improve osteoblast proliferation and differentiation in MC3T3-E1 mouse cells. The chemicals utilized in the experiment included: YK11, DHT, hydroxyflutamide (HF), ascorbic acid, and beta-glycerol phosphate; all obtained from Wako Pure Chemical Industries, Ltd. in Osaka, Japan. The MC3T3-E1 cells were cultured in a mixture of Minimum Essential Medium (MEM) alpha, and 10% fetal bovine serum (FBS) in a humidified 5% CO2 climate. After the cells were cultured, they were seeded on plates and maintained in a solution of MEM alpha and 10% charcoal-stripped FBS for 24 hours. In order to induce osteoblast differentiation, YK11 or DHT were added to the solution of MEM alpha, 10% csFBS, 50 ug/mL ascorbic acid, and 5 mM beta-glycerol phosphate [2].
MC3T3-E1 cells were seeded in 96-well microplates at a density of 5000 cells per well, followed by maintenance with 10% csFBS for 24 hours prior to treatment with YK11 or DHT. After 24 hours of treatment the cells were incubated for 96 hours and an MTS assay kit was used to evaluate the level of cell viability in the samples. The research team also investigated any changes in mineralization through the use of alizarin red S staining. Prior to staining, the cells were fixed with cold methanol for 20 min and rinsed three times with phosphate buffered saline (PBS). On day 10 the cells were rinsed three times with water in order to remove all traces of methanol. After the final rinse the cells were stained with 40 mM concentration of pH 6.38 alizarin red S stain solution for 15 minutes. In order to remove all unbound alizarin red S, the stained cells were rinsed with water 5 times. This process had to be completed before any culture images were taken [2].
Next, levels of alkaline phosphatase (ALP) activity through the use of Lab Assay ALP. The medium was removed at the experimental endpoints while the cell layers were rinsed three times with PBS and lysed by 0.05% TritonX-100. The remaining lysates were mixed with a 0.1 mM carbonate buffer that was composed of a solution of 2 nM MgCl2 and 6.7 mM of p-nitrophenyl phosphate, and followed by 15 minutes of incubation. NaOH was added to the mixture by the researchers in order to stop the reaction. Additionally, a real-time quantitative PCR (RT-qPCR) was completed by isolating total RNA. The RT-qPRC was conducted according to the manufacturer’s protocol using a 25 uL concentration of KOD SYBR qPCR Mix while results were analyzed using SDS software.
For further analytical purposes, cells were harvested and lysed in an SDS sample buffer that was composed of 123 mM Tris-HCl, pH 6.8, 4% SDS, 10% sucrose, 10 mM dithiothreitol, and 0.01% bromophenol blue. Any whole cell lysates were resolved by SDS-PAGE technology, while immunoblotting was completed using anti-Akt, anti-phospho Ser 473 Akt , and horseradish peroxidase-conjugated anti-tubulin antibody, as the primary antibodies. The research team used horseradish peroxidase-conjugated anti-mouse or rabbit immunoglobulin G antibody as the secondary antibody in the study, while band density was measured through the use of Image J software. All statistical analyses were performed via one-way ANOVA testing and Dunnett’s multiple comparison post-hoc test [2].
Discussion
1) As it was outlined in the selected data portion, the AR-overexpressing HEK193 cell line, named AR293, was used to examine the effects of YK11 on AR expressions. AR expression was not observed in the parent HEK293 cell line, but was able to be identified in 293AR cells. That being said, DHT and YK11 were used to treat the 293AR cell line due to their ability to activate the transcription process of the ARE-luciferase reporter gene. Results reported that ARE-luciferase reporter activity was elicited by both DHT and YK11 in the sub-micromolar range, however, it is important to note that the maximal activity of YK11 only reached approximately 10-20% of the maximal activity induced by DHT. When examining the effects of YK11 in the presence of DHT, 5 and 10 uM concentrations of YK11 were found to inhibit DHT-induced reported activity. Since DHT-induced activity was blocked by the YK11 expression, the researchers were able to conclude that the SARM successfully acts as a partial agonist of the androgen receptors [1].
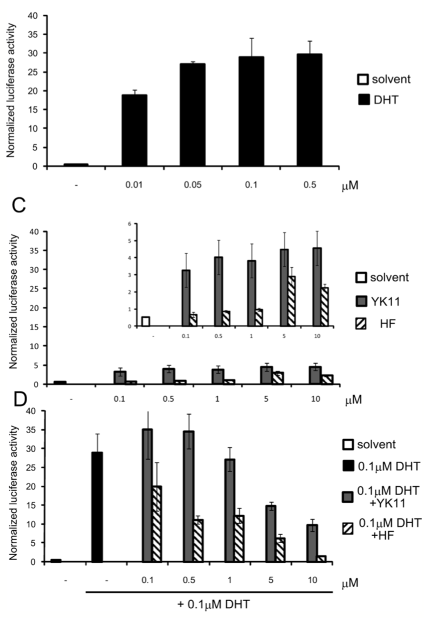
Figure 1: Effects of YK11 and DHT on ARE-luciferase activity
Furthermore, the research team examined how expression of AT-target genes responded to treatment with YK11 in the 293AR cells. Following 24 hours of treatment with 0.1 uM concentration of DHT or doses ranging from 0.1 to 10 uM of YK11, the RT-PCR found that FKBP51 mRNA expression was significantly enhanced with all treatment doses of YK11 to the same extent as 0.1 uM DHT. However, the mRNA expression did not show significant signs of improvement in the parent HEK293 cells. After concluding that improvement in FKBP51 and FGF18 mRNA expression was induced by both YK11 and DHT to the same levels, the next step of the study was to examine the effects of YK11 in the endogenous AR-expressing MD-MB 453 cells.
The previous results are in comparison to specifically androgen-regulated genes (SARG) mRNA expression; expression was only induced by DHT and not YK11. Induction of HSD11B2 mRNA expression elicited by DHT was stronger than mRNA expression elicited by YK11. It was also reported that the AR antagonist, bicalutamide (BIC), inhibited the induction of FKBP51 mRNA expression by YK11. This suggests that the ability of YK11 to affect gene expression is relative to the androgen agonist and dependent on the context of the gene [1].
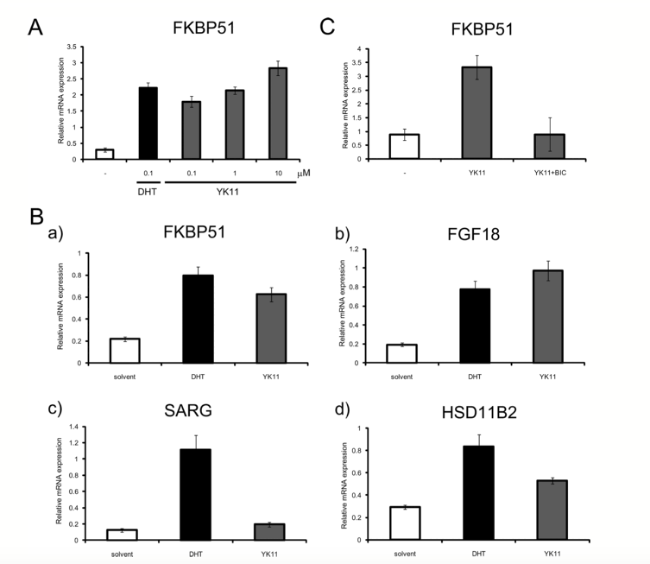
Figure 2: Changes in mRNA expression in response to treatment with YK11 versus DHT.
Results of the study were able to determine that the AR is localized in the cytoplasmic region in the absence of additional ligands. That being said, nuclear translocation is considered the first step of ligand-dependent activation of the AR. HEK293 cells were transfected with YFP-AR expressing plasmid that is localized in the cytoplasmic region in the absence of ligands. Additionally, YFP-AR tends to accumulate in the nucleus when both DHT and YK11 were present. This indicates nuclear translocation of the androgen receptor is accelerated by treatment with YK11.
In comparison to other nuclear receptors, androgen receptors are maximally active when a ligand is able to promote the interaction between the amino- and carboxy-terminal domains; this is referred to as an N/C interaction. A mammalian two-hybrid assay was completed during this study in order to observe the effects of YK11 on the N/C interactions. The assay reported when DHT is present, expression of fusion proteins GAL4-AR LBD and VP16-AR NTD resulted in a strong transactivation of the GAL4-reporter gene. The same results were not seen when YK11 was present; the SARM did not support transactivation of the reporter gene, but rather inhibit DHT-dependent transactivation of the reporter gene. These findings suggest that YK11 prevents DHT-induced N/C interactions between the LBD/AF-2 and NTD of AR in a similar manner to the DHT antagonist, hydroxyflutamide. The research team also determined that YK11 can activate the androgen receptor without causing an N/C interaction, supporting the hypothesis that YK11 acts as a partial agonist to the androgen receptor [1].
Overall, the researchers were able to conclude that in the presence of YK11, the two transactivation domains of the androgen receptor are able to work independently since there is no occurrence of an N/C interaction. The effects of YK11 on the transactivation domains were both assessed individually, revealing the expression plasmids coded for two AR mutants truncated at both ends: NTD-DBD and DBD-LBD. HEK293 cells were then transfected with the expression plasmids in combination with the ARE-luciferase reporter plasmid. Transfection of the HEK293 cells found that ligand-induced transactivation of the reporter gene was only observed in the presence of the full length AR [1].
2) An MTS assay was completed by researchers Yatsu et. Al in order to examine how YK11 affects osteoblast differentiation and cell proliferation. The MC3T3-E1 cells were treated with 0.5 uM of YK11 and 0.01 uM of DHT, over the course of 96 hours. Results of the treatment reported that both YK11 and DHT increased the rate of cell growth in the MC3T3-E1 cells. It was also found that these effects could easily be reversed by co-treatment with the AR antagonist, HF. These findings suggest that YK11 successfully accelerates osteoblast proliferation in a manner similar to DHT.
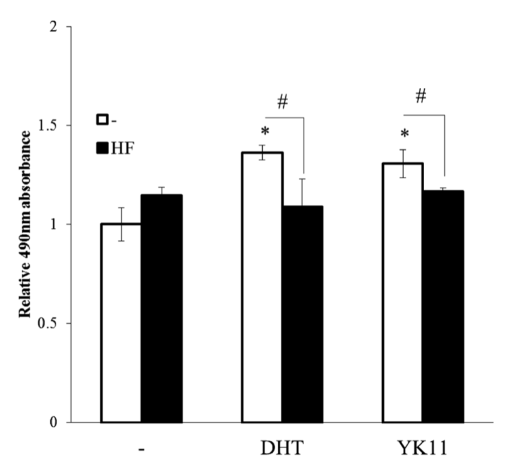
Figure 3: Effects of YK11 treatment on osteoblast proliferation
As the selected data mentioned, ALP activity was recorded since the enzyme acts as an early indicator of osteoblast differentiation. Over an experimental period of 10 days, the cells were cultured in the standard differentiation medium and treated with either YK11 or DHT. Both treatments led to an increased ALP activity in comparison to the cells treated with a control solvent. It is important to note that when the MC3T3-E1 cells were co-treated with AR antagonist, HF, ALP activity levels exhibited no significant increases [2].
Furthermore, calcium deposition in osteoblasts was identified through alizarin red S staining. The cells were cultured in the standard differentiation medium and treated with YK11 or DHT over a 3 week period of time. In comparison to the cells treated with a control solvent, mineralization was observed in the cells administered both the YK11 and DHT treatments. These results suggest that YK11 has the potential to promote osteoblasts differentiation through increased ALP activity and calcium deposition. Again, the effects elicited by YK11 were similar to those elicited by DHT.
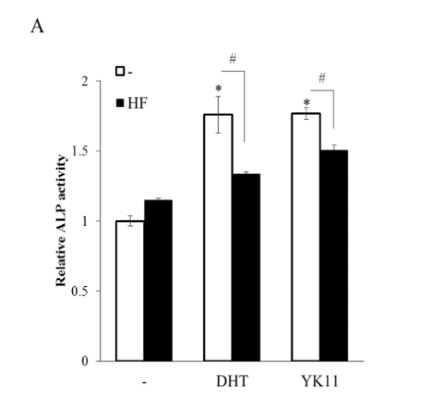
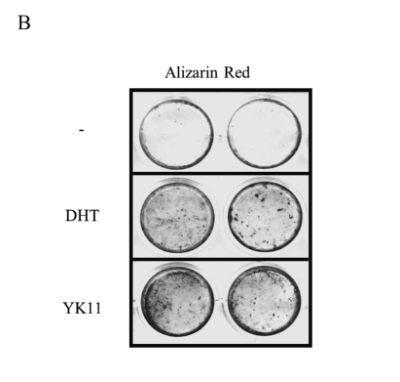
Figure 4: (A) Effects of YK11 treatment on osteoblast ALP activity (B) Effects of YK11 treatment on mineralization
The osteogenic marker osteoprotegerin (OPG) is an early indicator of osteoblast differentiation. On the other hand, levels of the osteogenic marker, osteocalcin (OC), indicate osteoblast differentiation later in the experimentation process. RT-qPCR was used in the cells treated with YK11 to measure mRNA expression; by day 4 of treatment YK11 increased OPG mRNA expression in a manner similar to DHT, the similarities between the effects elicited by the two treatments were still observed at day 14 of treatment.
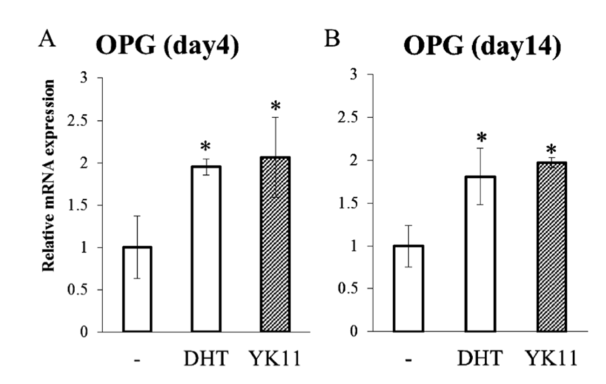
Figure 5: OPG expression in response to control, YK11, and DHT treatments
In terms of OC mRNA expression, no similarities were observed between treatment with YK11 versus DHT on day 4. However, by day 14 the two treatments exhibited similar elicited results. By day 14, the research team was able to observe dose-dependent increases on OC mRNA expression elicited by YK11 treatment. Similar to the previous differentiation tests, co-treatment with HF led to the inhibition of both YK11 and DHT. Overall, the findings of this study allowed the researchers to conclude that YK11 is able to promote osteoblast differentiation at a level similar to DHT in the MC3T3-E1 cells [2].
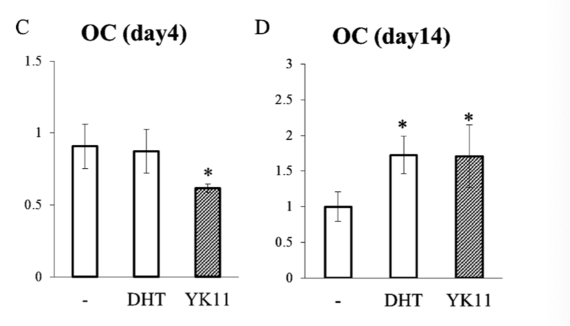
Figure 6: OC expression in response to control, YK11, and DHT treatments
The final experimental step was to evaluate the effects of YK11 on non-genomic signaling in the cells. Results of the examination initially reported that the androgen receptor was able to directly activate the PI3K/Akt pathway. It is important to mention that the PI3K/Akt pathway plays a crucial role in the regulation of osteoblast proliferation and differentiation. After 15 minutes of treatment with YK11 or DHT, the research team was able to determine the effects of YK11 on Akt signaling pathway activation. Similar to DHT, phosphorylation of the Akt protein was significantly enhanced in response to treatment with YK11. These findings suggest that YK11 has the potential to activate the Akt signaling pathway through non-genomic signaling in a manner similar to DHT [2].
Disclaimer
**LAB USE ONLY**
*This information is for educational purposes only and does not constitute medical advice. THE PRODUCTS DESCRIBED HEREIN ARE FOR RESEARCH USE ONLY. All clinical research must be conducted with oversight from the appropriate Institutional Review Board (IRB). All preclinical research must be conducted with oversight from the appropriate Institutional Animal Care and Use Committee (IACUC) following the guidelines of the Animal Welfare Act (AWA).
Citations
[1] Kanno Y, Hikosaka R, Zhang SY, Inoue Y, Nakahama T, Kato K, Yamaguchi A, Tominaga N, Kohra S, Arizono K, Inouye Y. (17α,20E)-17,20-[(1-methoxyethylidene)bis(oxy)]-3-oxo-19-norpregna-4,20-diene-21-carboxylic acid methyl ester (YK11) is a partial agonist of the androgen receptor. Biol Pharm Bull. 2011;34(3):318-23. doi: 10.1248/bpb.34.318. PMID: 21372378.
[2] Yatsu T, Kusakabe T, Kato K, Inouye Y, Nemoto K, Kanno Y. Selective Androgen Receptor Modulator, YK11, Up-Regulates Osteoblastic Proliferation and Differentiation in MC3T3-E1 Cells. Biol Pharm Bull. 2018;41(3):394-398. doi: 10.1248/bpb.b17-00748. PMID: 29491216.



| File Name | View/Download |
| 02-14-2023-Umbrella-Labs-YK-11-Certificate-Of-Analysis-COA.pdf |
VIEW CERTIFICATES OF ANALYSIS (COA)
Additional information
| Weight | 2 oz |
|---|---|
| Dimensions | 3 × 3 × 5 in |
| ChemSpider ID | |
| PubChem CID | |
| CAS Number |







