








SR-9009 STENABOLIC POWDER – 1000MG / 1 GRAM


$70.99
SR-9009 is sold for laboratory research use only. Terms of sale apply. Not for human consumption, nor medical, veterinary, or household uses. Please familiarize yourself with our Terms & Conditions prior to ordering.

*Includes:
- One (~10mg – 15mg) Red Micro Scoop
Also Available In:
![]() Liquid Option >>
Liquid Option >>
![]() Gel Option >>
Gel Option >>
- Description
- Additional information
Description
SR-9009 Stenabolic Powder
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
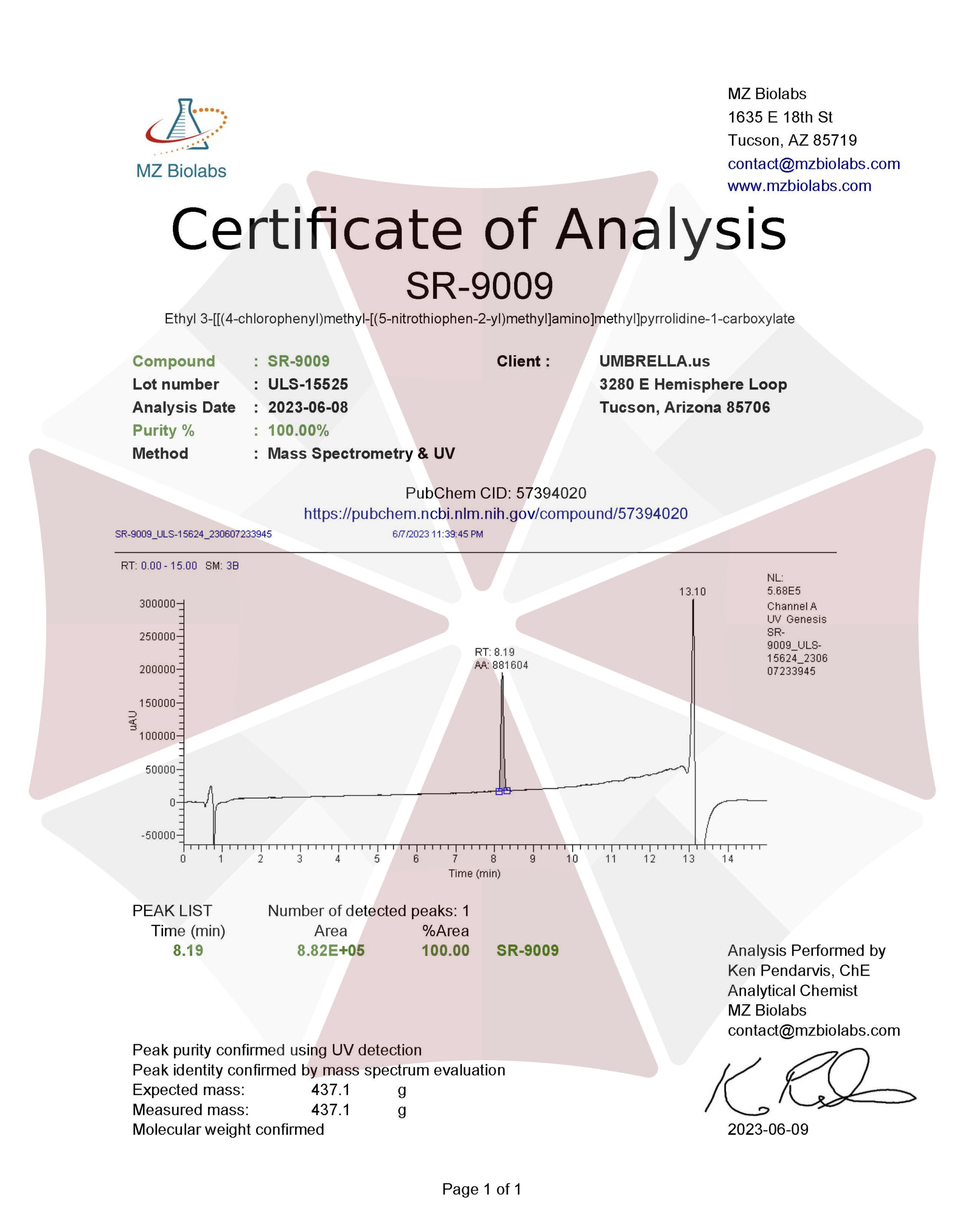
| CAS Number | 1379686-30-2 |
| Other Names | SR9009, SR 9009, Stenabolic, X5DCA09N30, CHEMBL1961796, REV-ERB Agonist II, UNII-X5DCA09N30, GTPL8901 |
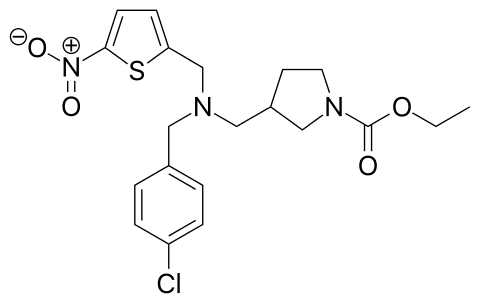
| IUPAC Name | ethyl 3-[[(4-chlorophenyl)methyl-[(5-nitrothiophen-2-yl)methyl]amino]methyl]pyrrolidine-1-carboxylate |
| Molecular Formula | C₂₀H₂₄ClN₃O₄S |
| Molecular Weight | 437.9 |
| Purity | ≥99% Pure (LC-MS) |
| Liquid Availability |
|
| Powder Availability | |
| Gel Availability | |
| Storage | Store in cool dry environment, away from direct sunlight. |
| Terms | All products are for laboratory developmental research USE ONLY. Products are not for human consumption. |
*Includes:
- One (~10mg – 15mg) Red Micro Scoop

What is SR-9009?
SR-9009 is a stenabolic SARM that is also categorized as a REV-ERB-A protein, and is typically found in the liver, skeletal muscles, and brain. The compound is best known for mediating circadian rhythm and metabolic activity. Many animal based studies have examined the potential of SR-900 to increase running speed, decrease levels of obesity, and stimulate mitochondrial production, thus enhancing muscle mass and metabolism.
Main Research Findings
1) As an REV-ERB agonist, SR-9009 has shown the potential to elicit anti-small-cell lung cancer effects through the inhibition of autophagy.
2) SR-9009 regulates proliferation, outgrowth, and suppression of neurons in a concentration-dependent manner.
Selected Data
1) The research team of Shen et. Al examined the ability of SR-9009 to prevent small-cell lung cancer through the inhibition of autophagy in a REV-ERB dependent manner. Natural circadian rhythm is able to coordinate cell proliferation and metabolism, as well as progress diseases like cancer. Due to the active role SR-9009 plays in mediation of the circadian clock, this experiment was conducted in order to determine the potential antitumor mechanism of the compound. The initial steps of the study investigated whether SR-9009 exhibited cytotoxicity to H69 and H446 chemosensitive cells as well as the corresponding chemoresistant cells, H69AR and H446DDP, these were labeled as small-cell lung cancer (SCLC) cells. The SCLC cells were treated with increasing concentrations over the course of 72 hours in order to determine the effects elicited by SR-9009. The research team also evaluated how the REV-ERBagonist affects cell migration and invasion as well as which characteristics were significant to the progression of cancer [1].
Typical antitumor drugs induce cell death through apoptosis. This led the researchers to question whether SR-9009 can promote apoptosis in SCLC cells. Western blot analysis was used to identify the presence of apoptotic proteins and changes in SCLC cell viability in response to treatment with SR-9009. Additionally, the research team used a general caspase family inhibitor to determine with apoptosis induced by SR-9009 was caspase-dependent. Subcutaneous tumor models were also established in mice with both chemosensitive and chemoresistant SCLC cells. The tumor-bearing mice were administered either SR-9009, chemotherapeutic drugs, or a combination of both.
The next portion of the experiment attempted to ascertain whether the effects of SR-9009 in SCLC cells are mediated by two different REV-ERB compounds, labeled as alpha and beta. Basal expression of REV-ERB-alpha and REV-ERB-beta was examined using human bronchial epithelial (HBE) cells as a control group, compared to the chemosensitive and chemoresistant SCLC cell lines. The results led the researchers to further examine whether expression of REV-ERBalpha can be increased with treatment with SR-9009. In order to specify the mechanism through which the SARM induces antitumor effects, small interfering RNA (siRNA) was used to known down REV-ERB-alpha [1].
Furthermore, examination of the effects of REV-ERB-alpha on the core autophagy gene, Atg5. Initial results were reported through the use of ChIP sequencing assays that reveal the presence of core autophagy genes. SR-9009 was then applied in order to identify the autophagy genes exhibiting repressed expression in response to REV-ERB-alpha binding to the promoter. Immunohistochemistry was also utilized to examine the expression of Atg5 in previously established SCLC subcutaneous tumor models. Activity of REV-ERB-alpha binding in the Atg5 promoter region was predicted through the UCSC database and analysis of Atg5 peaks in the initial ChIP-sequence assay. This procedure was followed by ChIP quantitative PCR assays.
The final step of the study was to further verify the autophagy-inhibiting effects of SRR-9009 by measuring autophagy activity in H69ARR and H466DDP cells that had been treated with SR-9009. Autophagosomes were evaluated through the use of laser confocal microscopy, TEM, and western blotting analysis [1].
2) Researcher Koli Shimozaki examined the effects elicited by SR-9009 stimulation of rat adult hippocampal neural stem/progenitor cells (AHPs). The rat AHPs were obtained from the Salk Institute and maintained in an undifferentiated state by culturing in an N2-medium containing DMEM, F12 medium supplemented with N2, and Invitrogen, as well as 20 ng/ml of FGF2. After 24 hours the undifferentiated maintenance medium was replaced with a neural differentiation medium containing 1 uM retinoic acid and 0.5% FBS, and was cultured for 7 days in order to induce neurogenesis in the AHPs. When inducing neural differentiation in the gene knockdown systems, the rat AHPs were transfected with knockdown plasmid vectors followed by washing three times with N2-medium and resuspension in neural differentiation medium before culturing on pO/L plates for 7 days [2].
DMSO was used to prepare the SR-9009 stock solutions; various concentrations of the SARM were used while the ratio of 1% DMSO and 50% ethanol was adjusted accordingly. The vehicle solution used as a control treatment was composed of 1% DMSO and 50% ethanol. Both the experimental and control treatments were diluted 200-fold and added into each culture system in a single administration. For the proliferation assay, the rat AHPs were left to incubate in the treatments for 24 hours and for the neurite outgrowth assay the samples were left for 7 days. Additionally, SR-9009 was added to the medium the day after the rat AHPs were transfected with plasmid DNA expressing shRNA targeting Rev-erb-beta. The cells were then cultured for an additional 24 hours to measure cell proliferation.
The next step of the study was to determine the functional relationship between SR-9009 and REV-ERB-beta. Rev-erb-beta mRNA was knocked down using a shRNA expression system while enhanced green fluorescent protein (GFP) was co-expressed under the control of the PGK promoter in order to successfully label transfected knockdown cells. The rats’ AHPs HCN1706 subline was transfected with plasmid DNA expressing scrambled shRNA or shRNA targeted against the Rev-erb-beta sequence through the use of the Nucleofector system. An RNeasy kit was used in order to extract total RNA from HCN1706 cells, while cDNA was transcribed using the ReverTra Ace Kit. An SYBR Green Q-PCR analysis kit allowed Shimozaki to conduct a qRT-PCR which was further examined with a QuantStudio 12 K Flex quantitative PCR system [2].
Both undifferentiated and differentiated rat AHPs were fixed in a 4% PFA solution and blocked using PBS containing 0.1% Triton X-100 and 1% donkey serum. Immunostaining was then performed using the primary antibodies: anti-NR1D2, anti-Nestin, aunty-Ki67, and anti-beta-III tubulin; as well as the secondary antibodies anti-rat, rabbit, or chicken DyLight 488, 549, and 649. Furthermore, DNA was stained using 4′-6-diamidino-2-phenylindole to identify cell nuclei. BrdU solution was added to rat AHPs in order to label proliferating cells. Prior to immunofluorescent staining with anti-BrdU antibody, the BrdU-treated cells were fixed with 4% PFA following treatment with hydrochloric acid. Detecting cells co-stained with anti-GFP and anti-BrdU antibodies were also used by Shimozaki to observe and measure proliferation rate of the cells [2].
Immunostaining images were taken using a confocal laser microscope and a fluorescence microscope while proliferation rate of the knockdown cells was measured by calculating the number of GFP-positive and GFP- and anti-BrdU double-positive cells stained with DAPI. This same procedure was followed in the samples treated with SR-9009. When observing the effects elicited by SR-9009 on neurite outgrowth, HC1706 cells were treated with varying concentrations of SR-9009, followed by immunostaining with anti beta-III tubulin antibodies and DAPI. Beta-III-tubulin-positive dendrites were measured using a BZ-X Analyzed and DP2-BSW application software. After determining the number of DAPI-positive cells, the average neurite length was found by dividing the total traced neurite length by the number of DAPI-positive cells. Additionally, Shimozaki counted the number of all neurite branching points and primary neurites that were formed directly from the cell bodies [2].
Discussion
1) When investigating the cytotoxic effects of SR-9009 on SCLC, initial results reported that cytotoxicity was induced by SR-9009 in a dose-dependent manner in both chemosensitive and chemoresistant cells, H69 and H446, and H69AR and H446DDP, respectively. This suggests that SR-9009 could potentially act as an effective treatment for first- or second-line treatment of SCLC. Wound healing assays and transwell migration assays emphasized the ability of SR-9009 to significantly decrease cell migratory behaviors, as well as inhibit invasive cellular behaviors [1].
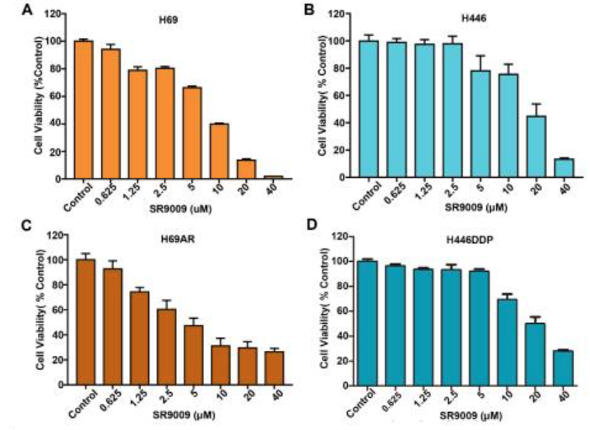
Figure 1: Cell viability in A) H69 chemosensitive cells, B) H446 chemosensitive cells, C) H69AAR chemoresistant cells, and D) H446DDP chemoresistant cells, in response to SR-9009
Additionally, western blot assays were used to observe the activation of apoptotic proteins in response to SR-9009 treatment; the proteins identified were PARP and caspase 3. Immunofluorescence assays for cleaved caspase 3 and TUNEL assays revealed that SR-9009 impaired SCLC cell viability, partially due to the induction of apoptosis. When determining whether apoptosis induced by SR-9009 was caspase-dependent the research team found that following pretreatment with z-VAD-fmk, cleaved caspase 3 and PARP were significantly decreased. However, results also revealed that SR-9009-induced cytotoxicity could be partially restored by z-VAD-fmk. These results allowed the researchers to conclude that SR-9009 has the potential to enhance caspase-dependent apoptosis in SCLC cells [1].
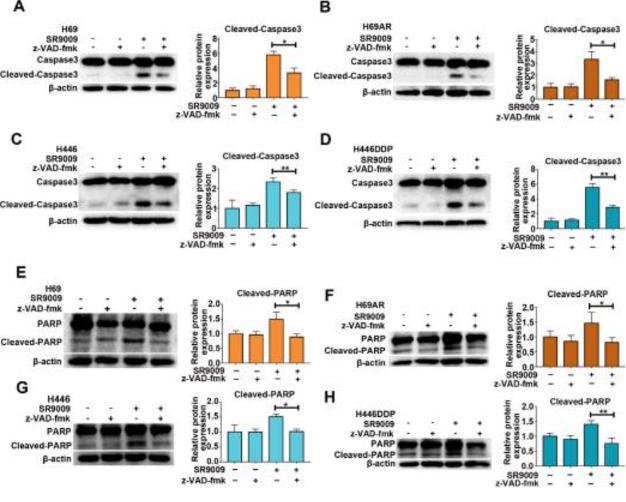
Figure 2: changes in caspase 3 and PARP in response to SR-9009 treatment in SCLC cells.
For the second portion of the study subcutaneous tumor models were established in mice with both types of SCLC cells. Results found that treatment with SR-9009 led to an inhibition of tumor growth in both the chemosensitive and chemoresistant models. Additionally, pretreatment with SR-9009 showed the potential to effectively sensitize resistant cells to chemotherapy. The data collected allowed the research team to conclude that SR-9009 exhibits potent antitumor effects against SCLC, independent of the chemoresistant status of the cells.
Further analysis occurred in order to determine whether the effects of SR-9009 are dependent on the expression of the REV-ERB proteins. Compared to HBE control cells, SCLC cells expressed high levels of REV-ERB-alpha and low levels of REV-ERB-beta, however protein expression of REV-ERB-alpha was increased after administration of SR-9009. When REV-ERB-alpha was knocked down by siRNA, the downregulation of the protein resulted in the inhibition of the anticancer activity of SR-9009. This indicated that REV-ERB-alpha is related to the antitumor effects of SR-9009 in SCLC cells [1].
The research team used ChIP-sequencing assay to determine the mechanism of action of SR-9009 in SCLC cells. Results of the assay revealed the presence of peaks in the core autophagy genes Atg5, Atg2A, Atg4C, Atg7, Atg13, Atg16L1, Atg101, AMBRA1, DRAM2, and Ulk1. This suggests that the REV-ERB-alpha compound can potentially bind to the promoters of autophagy genes in SCLC cells, leading to repressed expression of these genes. SR-9009 was then applied to emphasize which autophagy genes experienced repressed expression. The Atg5 gene was most significantly blocked following treatment with SR-9009 in both chemoresistant cell types. Immunohistochemistry solidified the initial findings, highlighting the significant inhibition in Atg5 gene expression in the SR-9009 treatment group [1].
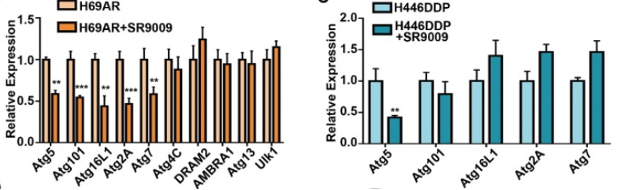
Figure 3: Relative expression in the chemoresistant cells with and without SR-9009 treatment
ChIP-qPCR assays were conducted next in order to further investigate the role REV-ERB-alpha plays in the regulation of the autophagy gene, Atg5. Results of the assay found that REV-ERB-alpha was enriched in the Atg5 promoter. This indicates that Atg5 expression could potentially be repressed by the binding of REV-ERB-alpha to the Atg5 promoter, thus inhibiting promoter activity. The research team was able to conclude from the data that the Atg5 gene is a direct target of REV-ERB-alpha, and that the protein negatively regulates the autophagy gene in SCLC cells [1].
2) The first portion of the study was to analyze the expression and subcellular localization of REV-ERB-beta in rat AHP. Shimozaki was able to confirm that the REV-ERB-beta protein is expressed and localized in the nucleus and cytoplasm of all cultured rat AHPs. The cultured rat AHPs uniformly expressed the intermediate filament Nestin; Nestin acts as an undifferentiated neural stem/progenitor cell marker protein. Ki67, the nuclear protein and cell proliferation maker, was also detected in cultured AHPs. A similar pattern of expression is identified in cultured neural stem/progenitor cells derived from adult mouse brains. These results suggest that there is a common expression pattern for REV-ERB-beta, specifically in rodents. This data indicates the cell-cycle dynamics are not linked to the localization of REV-ERB-beta [2].
Neuronal and glial cell differentiation was induced in cultured rat AHPs, in order to analyze the expression and subcellular localization of REV-ERB-beta during adult brain neurogenesis. The rate of neurogenesis in the hippocampal dentate gyrus of the adult mouse brain was reported to be approximately 70%, while the HCN1706 cell line used in this study exhibited a neuronal differentiation rate of more than 90%.
Furthermore, the reported results of immunostaining the cultured rat AHPs suggested that the REV-ERB-beta protein was more strongly localized in the nucleus under neural differentiation conditions rather than non-differentiating conditions. Rev-erb-beta expression in cultured rat AHPs was also compared in neural differentiation versus non-differentiating conditions. Results reported that under neuronal differentiation conditions Rev-erb-beta mRNA expression was more than two-fold that of the samples under non-differentiating conditions. Overall, these research findings suggest that REV-ERB-beta accumulated in the nucleus and expression of the protein was enhanced by the induction of neurogenesis [2].
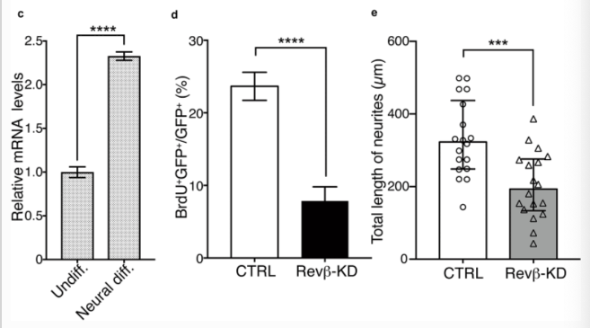
Figure 4: changes in C) Rev-erb-beta expression levels, D) percent of BrdU/GFP expression, E) total length of neurites.
Proliferation in Rev-erb-beta specific knockout samples was measured by adding BrdU to the medium. The collected data found that the knockdown of Rev-erb-beta in AHPs significantly reduced BrdU uptake in comparison to the control group. This indicates that proliferation in rat AHPs is dependent on the presence of Rev-erb-beta.
When investigating the relationship between neurite outgrowth and REV-ERB-beta, the medium was replaced with a neural differentiation medium following electroporation followed by the induction of neural differentiation for 7 days. Anti-beta-III-tubulin antibody immunostaining revealed the molecular and physical properties of the differentiated AHPs by measuring the neurite projection lengths of GFP-positive cells. Results of this analysis found that neurite growth was significantly diminished in Rev-erb-beta knockdown cells in comparison to control cells. These findings suggest that REV-ERB-beta participates in the process of proliferation and neurite outgrowth in cultured rat AHPs in a manner similar to the reported results for the cultured adult mouse neural stem/progenitor cells [2].
Shimozaki observed changes in proliferation and neuronal differentiation in cultured rat AHPs after adding various concentrations of SR-9009 to the medium in order to further characterize the cellular properties associated with enhancement of REV-ERBs in response to activation with SR-9009. Proliferation was measured through the BrdU uptake method; treatment with a vehicle compound did not affect proliferation in rat AHPs. There were no significant changes in BrdU uptake when SR-9009 was administered at concentrations between 0 and 1 uM, however, when SR-9009 was added at concentrations about 1.5 uM, decreased proliferation was detected; equilibrium was reached at 2.5 uM. This data suggests that the growth arrest signal could potentially be activated by SR-9009 when the concentration exceeded a specific amount.
The next step of the study was to examine how knock down Rev-erb-beta in cultured rat AHPs determine the REV-ERB-beta dependence of effects mediated by SR-9009. When the cultured AHPs were transfected with a control vector, a reduced rate in BrdU uptake was observed after 2.5 uM of SR-9009 was added to the samples. It is important to note that BrdU uptake was significantly reduced after SR-9009 was added, even with the Rev-erb-beta gene being knocked down. These findings allowed Shimozaki to conclude that both REV-ERB-beta-dependent and -independent pathways are involved in the SR-9009-mediated inhibition of AHP proliferation [2].
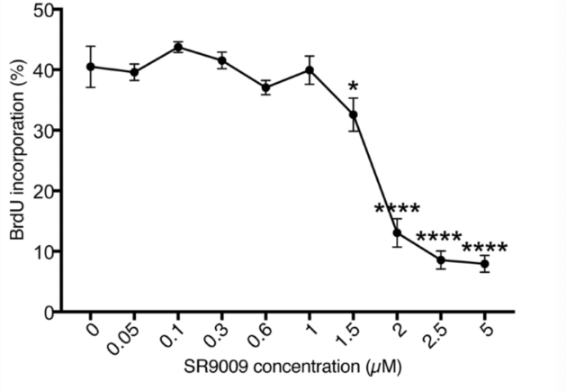
Figure 5: Changes in BrdU uptake in response to varying concentrations of SR-9009.
When examining neurite outgrowth during neurogenesis, dendritic outgrowth was induced when concentrations between 0 and 1 uM of SR-9009 were added to the medium. Maximum growth was observed at a concentration of 0.1 um of SR-9009. However, dendritic growth was suppressed in concentrations higher than 2.5 uM, indicating that SR-9009 induces positive and negative effects on neurite outgrowth during neurogenesis. Cultured cells were fixed with 4% PFA and immunostained to reveal that cells transfected with control shRNA resulted in increased neurite length in comparison to the untreated cells. When the Rev-erb-beta gene was knocked down, the average neurite length of differentiated AHPs did not increase with the addition of low-concentrations (0.1 uM) of SR-9009. Addition of high-concentration SR-9009 (2.5 uM) to control cells resulted in a decrease in the mean neurite length of differentiated AHPs. However when SR-9009 was added to cells with a knocked down Rev-erb-beta gene, no suppressive activity was identified [2].
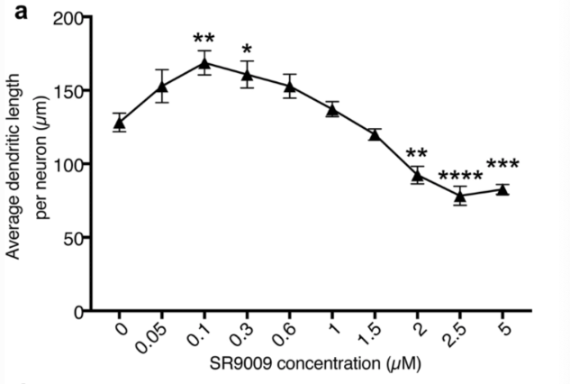
Figure 6: Changes in dendritic length in response to varying concentrations of SR-9009
The results of the study solidified the hypothesis that SR-9009 reduces proliferation in rat AHPs and induces positive and negative effects on neurite outgrowth. Shimozaki attempted to identify the underlying mechanism of action, by examining whether the expression levels of the Rev-erb-beta gene and REV-ERB-beta target genes were affected by SR-9009. The qRT-PCR conducted found that adding high-concentration of SR-9009 did not lead to any dramatic chances in Rev-erb-beta mRNA expression, however, expression of the REV-ERB-beta target gene Ccna2 was significantly reduced by high concentrations of SR-9009. After 7 days of culturing under neural differentiation conditions after treatment with low and high concentrations of SR-9009, there were no significant changes observed in the expression of Rev-erb-beta or the REV-ERB-beta target gene, Sez6, by either dose of the SARM. This indicates that SR-9009 did not affect the expression of Rev-erb-beta but rather the expression of various REV-ERB-beta target genes [2].
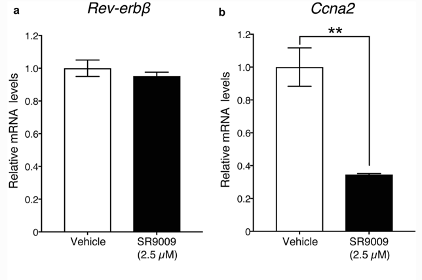
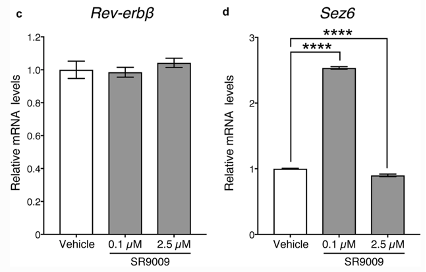
Figure 7: relative mRNA expression in Rev-erb-beta gene and REV-ERB-beta target genes, Ccna2 and Sez6
Disclaimer
**LAB USE ONLY**
*This information is for educational purposes only and does not constitute medical advice. THE PRODUCTS DESCRIBED HEREIN ARE FOR RESEARCH USE ONLY. All clinical research must be conducted with oversight from the appropriate Institutional Review Board (IRB). All preclinical research must be conducted with oversight from the appropriate Institutional Animal Care and Use Committee (IACUC) following the guidelines of the Animal Welfare Act (AWA).
Citations
[1] Shen W, Zhang W, Ye W, Wang H, Zhang Q, Shen J, Hong Q, Li X, Wen G, Wei T, Zhang J. SR9009 induces a REV-ERB dependent anti-small-cell lung cancer effect through inhibition of autophagy. Theranostics. 2020 Mar 15;10(10):4466-4480. doi: 10.7150/thno.42478. PMID: 32292508; PMCID: PMC7150483.
[2] Solt LA, Wang Y, Banerjee S, Hughes T, Kojetin DJ, Lundasen T, Shin Y, Liu J, Cameron MD, Noel R, Yoo SH, Takahashi JS, Butler AA, Kamenecka TM, Burris TP. Regulation of circadian behaviour and metabolism by synthetic REV-ERB agonists. Nature. 2012 Mar 29;485(7396):62-8. doi: 10.1038/nature11030. PMID: 22460951; PMCID: PMC3343186
SR-9009 is sold for laboratory research use only. Terms of sale apply. Not for human consumption, nor medical, veterinary, or household uses. Please familiarize yourself with our Terms & Conditions prior to ordering.



| File Name | View/Download |
| 06-08-2023-Umbrella-Labs-SR-9009-Stenabolic-Certificate-Of-Analysis-COA.pdf |
VIEW CERTIFICATES OF ANALYSIS (COA)
Additional information
| Weight | 2 oz |
|---|---|
| Dimensions | 3 × 3 × 5 in |
| CAS Number | |
| PubChem CID | |
| ChEMBL ID | |
| ChemSpider ID |







